 [English]
[English]
| [English]
| [note bibliographique] | [sommaire] |
L'auteur ťtudie le lancement de la tomate qui provoquait la rťaction yellante chez la cantatrice et dťmontre que plusieurs aires de la cervelle ťtaient impliquťes dans la rťponse, en particulier le trajet lťgumier, les nuclei thalameux et le fiÁure musicien de l'hťmisphŤre nord.*
Les effets frappants du jet de tomates sur les sopranos, observťs aux heures ultimes du siŤcle dernier par Marks et Spencer (1899) qui, les premiers, employŤrent le terme de rťaction de hurlements (RH), ont ťtť largement dťcrits dans la littťrature. Si de nombreuses ťtudes expťrimentales (Zeeg & Puss, 1931; Roux & Combaluzier, 1932; Sinon & coll., 1948), anatomopathologique (Hun & Deu, 1960), comparative (Karybb & Szyla, 1973) et prospective (Else & Vire, 1974) ont permis de dťcrire avec prťcision ces rťponses caractťristiques, les donnťes neuroanatomiques, aussi bien que neurophysiologiques sont, en dťpit de leur grand nombre, ťtonnamment confuses. Dans leurs dťmonstrations dťsormais classiques, publiťes dans la fin des annťes 20. Chou & Lai (1927 a, 16, c, 1928 a, 16, 1929 a, 1930) ont ťcartť l'hypothŤse d'un simple rťflexe nociceptif facio-facial qui avait ťtť ťmise il y a de nombreuses annťes par certains auteurs (Mace & Doyne, 1912; Payre & Tairnelle, 1916; Sornette & Billevayze, 1925).
Depuis lors, de nombreux travaux ont ťtť menťs pour tenter de rťsoudre l'ťnigme embrouillťe ainsi que l'embrouillement ťnigmatique des versants affťrent et/ou effťrent de la RH et ont conduit ŗ incriminer, de maniŤre quelque peu chaotique, une multitude de structures et de voies : l'existence de voies affťrentes trigťminale (Loewenstein et coll., 1930), bitrigťminale (Von Aitick, 1940), quadritrigťminale (Van der Deder, 1950), supra, infra et intertrigťminale (Mason & Ragoun, 1960) a ťtť invoquťe avec une certaine vraisemblance, de mÍme que celle de canaux d'entrťe maculaire (Zakouski, 1954), sacculaire (Bortsch, 1955), utriculaire (Malosol, 1956), ventriculaire (Tarama, 1957), monoculaire (Zubrowska, 1958), binoculaire (Chachlik, 1959-1960), trioculaire (Strogonoff, 1960), auditif (BalalaÔka, 1515) et digestif (Alka-Seltzer, 1815). Des circuits spinothalamiques (Attou & Ratathou, 1974), rubrospinal (Maotz & Toung, 1973), nigrostriť (Szentagothai, 1972), rťticulaire ( Pompeiano et coll. 1971), hypothalamique (Hubel & Wiesel, 1970), mťsolimbique (Kuffler, 1969) et cťrťbelleux (High & Low, 1968) ont ťtť explorťs en vain, pour tenter d'ťlucider l'organisation de la RH, et la responsabilitť de presque toutes les parties du cortex somesthťsique (Pericoloso & Sporghersi, 1973), moteur (Ford, 1930), comissural (Gordon & Bogen, 1974) et associatif (Einstein et coll., 1974) a ťtť ťvoquťe dans le dťveloppement progressif de la rťponse, bien que, jusqu'ŗ maintenant, les mťcanismes affťrents et effťrents de la RH n'aient jamais ťtť explicitťs de maniŤre dťcisive et convaincante.
Unsofort & Tchetera ont observť que " plus on jette de tomates sur les sopranos
et plus elles crient "; par ailleurs, des ťtudes comparatives, par rapport ŗ
la rťaction de gasp (Otis & Pifre, 1964), au hoquet (Carpentier & Fialip, 1964),
au ronronnement du chat (Remmers & Gautier, 1972), au rťflexe HM (Vincent et coll., 1976),
ŗ la ventriloquie (Mc Culloch et coll., 1964), aux cris perÁants, aigus ou stridents et
aux autres rťactions hystťriques (Sturm & Drang, 1973) provoquťes par le jet de tomates,
aussi bien que de choux, pommes, tartes ŗ la crŤme, chaussures, billots et enclumes
(Harvar & Mercy, 1973) ont conduit ŗ l'hypothŤse solide selon laquelle la RH est
dťterminťe par un mťcanisme de rťtroaction positive, qui repose sur une interdigitation
semilinťaire quadristable ŗ embranchements multiples de sous-rťseaux neuronaux fonctionnant
en dťsordre*. (Beulott et coll., 1974).
Bien que cette hypothŤse soit assez sťduisante, les donnťes anatomiques et physiologiques
dont nous disposons sont insuffisantes pour pouvoir l'ťtayer. Nous avons donc dťcidť
d'explorer de maniŤre systťmatique l'organisation interne croissante ou dťcroissante
de la RH pour tenter d'ťlaborer un modŤle anatomique.
Les donnťes ont ťtť contrŰlťes par rapport au lancement d'autres projectiles: trognons de pomme,
rognures de choux, chapeaux, roses, citrouilles, balles de fusil et ketchup (Heinz, 1952).
Enregistrement
L'activitť des diffťrentes aires cťrťbrales a ťtť enregistrťe par l'intermťdiaire de semi
macro-ťlectrodes en alliage verre-tungstŤne placťes au petit bonheur*
selon la mťthode de Zyszytrakyczywsz-Sckrawszhwez (1974). La dťtection des pointes a ťtť assurťe
par audiomonitoring : chaque fois qu'une dťcharge ťtait entendue, elle ťtait soigneusement
photographiťe, enregistrťe, affichťe sur un monographe et, aprŤs intťgration, sur un polygraphe.
L'analyse statistique des rťsultats a ťtť rťalisťe au moyen d'un algorithme inspirť du
tennis (Wimbledon, 1974), c'est-a-dire que chaque fois qu'une structure gagnait un jeu,
elle ťtait considťrťe comme ťtant impliquťe dans la RH.
| Tableau 1 : Rťponses des diffťrentes parties du cerveau
ŗ la stimulation tomatotopique ŗ diverses frťquences | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
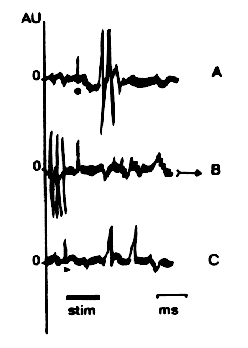
| Fig. 1 : Activitť des structures rťagissant ŗ la stimulation tomatique. Le trait horizontal indique le dťbut et la fin du stimulus. Etalonnage : 3,1416 ms. Chaque tracť est formť par la superposition de 33,57 enregistrements successifs. Remarquez le point en A, la flŤche en B et le triangle noir en C. |
Des exemples de rťponses de ces structures sont reprťsentťs sur la figure 1, oý l'analyse
temporelle de la distribution des pointes fondťe sur les caractťristiques de la rťpartition
alťatoire temporelle programťe (RATP) des aires ťtudiťes permet de distinguer 3 sous-types
d'unitťs cťrťbrales : 1 ) les unitťs rťpondant avant la stimulation ; 2 ) les unitťs rťpondant
au cours de la stimulation ; 3 ) les unitťs rťpondant aprŤs la stimulation.
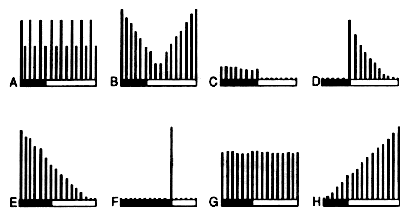
La comparaison des rťponses obtenues avec la stimulation par le ketchup et d'autres projectiles,
rťsumťe dans la figure 2, apporte des arguments indiscutables en faveur de l'existence d'une
organisation tomatotopique de la RH le long, entre et ŗ travers le NARTpl, la paTL et le scMS.
|
| Fig. 2 : Exemples de rťponses provoquťes par la tomate et d'autres projectiles. Explications dans le texte A = tomate, B = pomme, C = chou, D = chapeau, E= roses, F = ketchup®, G = potiron, H = balle de fusil |
|
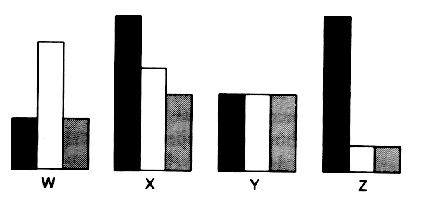
| Fig. 3 : Relation temporelle entre les rťponses enregistrťes dans la rťgion impliquťe dans la RH. Abcisse : unitťs arbitraires ; ordonnťes : unitťs internationales.Explications dans le texte |
|
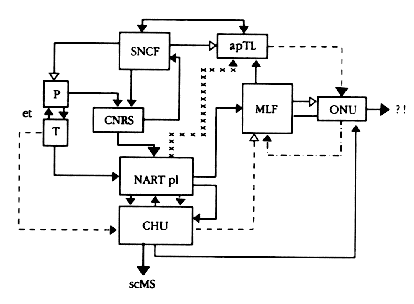
| Fig. 4 : Tentative d'ťlaboration d'un modŤle anatomique du mťcanisme de la RH. Explications dans le texte ou ailleurs. Lignes contin ues = inhibition, lignes discontinues = interrogation, lignes ťtoilťes = redhibition, lignes en pointillť = substentation. |
Ce travail a pu Ítre menť gr‚ce aux subventions accordťes par le syndicat rťgional
des producteurs de fruits et lťgumes,
l'association franÁaise des amateurs d'art lyrique (AFAAL)
et la fťdťration internationale des dactylo-bibliographes (FIDB).
|
Note du webmestre : Tous droits rťservťs sur ce texte. Georges Perec ťcrivit cette version franÁaise ŗ l'occasion du jubilťe de madame Bonvallť, directrice de recherche ŗ l'INSERM. Elle a ťtť publiťe dans le Jim (Journal International de Mťdecine) nį 103, rubrique "Pitres et Travaux" dans le milieu des annťes 1980. Mis en ligne avec l'aimable autorisation du Jim le 28 juil 2004. La version habituellement connue et que vous pouvez lire par le lien English est publiťe au SEUIL collection "La librairie du XXį siŤcle" dans le recueil "CANTATRIX SOPRANICA L. et autres ťcrits scientifiques" ...retour Je suis heureux que les deux versions puissent Ítre ainsi confrontťes... |
